Recombinant Mouse NF-kappa-B essential modulator (Ikbkg)
-
中文名稱:小鼠Ikbkg重組蛋白
-
貨號:CSB-YP011574MO
-
規格:
-
來源:Yeast
-
其他:
-
中文名稱:小鼠Ikbkg重組蛋白
-
貨號:CSB-EP011574MO
-
規格:
-
來源:E.coli
-
其他:
-
中文名稱:小鼠Ikbkg重組蛋白
-
貨號:CSB-EP011574MO-B
-
規格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:小鼠Ikbkg重組蛋白
-
貨號:CSB-BP011574MO
-
規格:
-
來源:Baculovirus
-
其他:
-
中文名稱:小鼠Ikbkg重組蛋白
-
貨號:CSB-MP011574MO
-
規格:
-
來源:Mammalian cell
-
其他:
產品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
別名:Ikbkg; Nemo; NF-kappa-B essential modulator; NEMO; IkB kinase-associated protein 1; IKKAP1; mFIP-3; Inhibitor of nuclear factor kappa-B kinase subunit gamma; I-kappa-B kinase subunit gamma; IKK-gamma; IKKG; IkB kinase subunit gamma; NF-kappa-B essential modifier
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Full length protein
-
表達區域:1-412
-
氨基酸序列MNKHPWKNQL SEMVQPSGGP AEDQDMLGEE SSLGKPAMLH LPSEQGTPET LQRCLEENQE LRDAIRQSNQ MLRERCEELL HFQVSQREEK EFLMCKFQEA RKLVERLSLE KLDLRSQREQ ALKELEQLKK CQQQMAEDKA SVKAQVTSLL GELQESQSRL EAATKDRQAL EGRIRAVSEQ VRQLESEREV LQQQHSVQVD QLRMQNQSVE AALRMERQAA SEEKRKLAQL QAAYHQLFQD YDSHIKSSKG MQLEDLRQQL QQAEEALVAK QELIDKLKEE AEQHKIVMET VPVLKAQADI YKADFQAERH AREKLVEKKE YLQEQLEQLQ REFNKLKVGC HESARIEDMR KRHVETPQPP LLPAPAHHSF HLALSNQRRS PPEEPPDFCC PKCQYQAPDM DTLQIHVMEC IE
-
蛋白標簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產品評價

靶點詳情
-
功能:Regulatory subunit of the IKK core complex which phosphorylates inhibitors of NF-kappa-B thus leading to the dissociation of the inhibitor/NF-kappa-B complex and ultimately the degradation of the inhibitor. Its binding to scaffolding polyubiquitin plays a key role in IKK activation by multiple signaling receptor pathways. Can recognize and bind both 'Lys-63'-linked and linear polyubiquitin upon cell stimulation, with a much highr affinity for linear polyubiquitin. Could be implicated in NF-kappa-B-mediated protection from cytokine toxicity. Essential for viral activation of IRF3. Involved in TLR3- and IFIH1-mediated antiviral innate response; this function requires 'Lys-27'-linked polyubiquitination.
-
基因功能參考文獻:
- NEMO deficiency hampered activation of IKK complex in osteoclast precursors, causing arrest of osteoclastogenesis and apoptosis. Interestingly, inhibiting apoptosis by genetic ablation of TNFr1 significantly increased cell survival, but failed to rescue osteoclastogenesis or reverse osteopetrosis. PMID: 27435916
- NEMO was critically involved in the cGAS-STING pathway. PMID: 28939760
- This study provides evidence for a protective function of canonical IKK/NF-kappaB signalling in the arenchymal compartment during pancreatitis. In the absence of the regulatory IKK subunit NEMO, a combination of a fibrogenic gene signature, enhanced acinar-ductal metaplasia and a lack of regeneration contribute to the exacerbation of pancreatitis. PMID: 27464707
- E+P treatment of breast cancer cells increased ER binding to the NEMO promoter, thereby increasing NEMO expression. PMID: 28515148
- robust caspase activation in NEMO-deficient cells is concomitant with RIPK3 recruitment to the apoptosis-mediating complex. PMID: 27560715
- Whereas Parkin has been reported to positively regulate the expression of OPA1 through NEMO, herein we found that PARK2 overexpression did not modify the expression of OPA1. PMID: 26024391
- prevents colon inflammation by NF-kappaB-independent functions PMID: 26982364
- Thyrocyte-specific NEMO knock-out mice gradually develop hypothyroidism after birth, which leads to reduced body weight and shortened life span. PMID: 26786105
- SENP1 deletion in adipocytes causes Type 1 diabetes mellitus via enhanced SUMOylation of NEMO, leading to increased NF-kappaB activity, cytokine production and pancreatic inflammation. PMID: 26596471
- Crucial functions of inflammatory TAK1-NEMO signaling in protecting the brain endothelium and maintaining normal brain function. PMID: 26347470
- Data show that NEMO protein prevents hepatocarcinogenesis by inhibiting receptor (TNFRSF)-interacting serine-threonine kinase 1 (RIPK1) kinase activity-driven hepatocyte apoptosis. PMID: 26555174
- Data show that the in utero death of NF-NF-kappaB essential modulator (NEMO) and cylindromatosis protein double mutant mice is mediated by TNF receptor 1 (TNFR1) signaling and can be rescued by TNFR1 deficiency. PMID: 26224629
- signaling through NEMO might not only be involved in the production of NF-kappaB proinflammatory chemokines but also regulates podocyte dynamics independently of NF-kappaB PMID: 26268269
- The stability of the NEMO coiled coil is maintained by strong interhelix interactions in the region centered on residue 54. PMID: 25400026
- NEMO deficiency results in apoptosis, the development of liver fibrosis and hepatocellular carcinoma, all of which are substantially alleviated by allogeneic hepatocyte transplantation. PMID: 24979756
- Mass spectrometric analysis demonstrated that WA covalently modifies NEMO on a cysteine residue within the C-terminal zinc finger (ZF) domain. Point mutations to the ZF can reverse the WA-induced Lys-48-polyubiquitin binding phenotype PMID: 25296760
- Authors propose a model in murine cytomegalovirus virion-associated protein M45 delivered by viral particles activates NF-kappaB, presumably involving an interaction with RIP1 and NEMO. PMID: 24942588
- NEMO ubiquitination is required for optimal innate immune signaling responses. These findings suggest that NEMO ubiquitination is crucial for NF-kappaB activity in response to innate immune agonists. PMID: 23871670
- NEMO ZF, like other NEMO related-ZFs, binds mono-Ub and di-Ub with distinct stoichiometries, indicating the presence of a new Ub site within the NEMO ZF. PMID: 24100029
- NEMO, an IKK subunit necessary for canonical NF-kappaB activation, is dispensable for normal pancreatic development and function, but essential for the propagation of KRAS-induced PanIN lesions. PMID: 22751123
- The E3 ligase parkin maintains mitochondrial integrity by increasing linear ubiquitination of NEMO PMID: 23453807
- In livers of mice, expression of NEMO and activation of NF-kappaB are required for hepatocyte proliferation and liver regeneration. PMID: 22922425
- NEMO binds ubiquitinated TANK-binding kinase 1 (TBK1) to regulate innate immune responses to RNA viruses. PMID: 23028469
- Murine cytomegalovirus inhibits the inflammatory cascade by blocking Toll-like receptor and IL-1 receptor-dependent NF-kappaB activation through an interaction of the viral M45 protein with the NF-kappaB essential modulator. PMID: 22319449
- When peptides containing the NF-kappaB essential modulator (NEMO) binding domain disrupt the IkappaB kinase (IKK)gamma complex, NF-kappaB activation is blocked, necrosis is decreased and regeneration is increased. PMID: 21624467
- NEMO Lysine392, a well-established K63-linked polyubiquitination site, is an important mediator of polymethylmethacrylate-induced osteolysis. PMID: 21913221
- tumor necrosis factor alpha-induced necroptosis requires the adaptor proteins FADD and NEMO. PMID: 21746883
- Data show that DNA damage stimulates the formation of a cytosolic complex of ATM, NEMO, RIP1, and TAK1, and that TAK1 mediates the NF-kappaB and p38 mitogen-activated protein kinase (MAPK)/MAPK-activated protein 2 responses to DNA damage. PMID: 21606198
- ANXA1 can constitutively activate NF-kappaB in breast cancer cells through the interaction with the IKK complex PMID: 21383699
- the effects of individually deleting the NEMO-binding domains in IKK alpha and IKK beta on classical NF-kappa B signaling and IKK complex formation. PMID: 19666475
- binding of linear ubiquitin chains to NEMO in NF-kappaB activation PMID: 21153314
- PAD2 interacts with IKKgamma and suppresses NF-kappaB activity. PMID: 20937835
- ATM- and NEMO-dependent ubiquitination of ELKS leads to the ubiquitin-dependent assembly of TAK1/TAB2/3 and NEMO/IKK complexes, resulting in IKK and NF-kappaB activation following genotoxic stimuli. PMID: 20932476
- RNAi down-regulation of the expression of IKKalpha and IKKgamma also down-regulated the expression of NF-KappaB p65, p50 and p105 proteins both in cytoplasm and nucleus. PMID: 19278579
- A mutation of Ikbkg causes immune deficiency without impairing degradation of IkappaB alpha. PMID: 20133626
- BAG3 alters the interaction between HSP70 and IKKgamma, increasing availability of IKKgamma and protecting it from proteasome-dependent degradation; this, in turn, results in increased NF-kappaB activity and survival PMID: 20368414
- the ADAP CARMA1 binding site is required for IKK gamma ubiquitination; both TAK1 and CARMA1 binding sites are required for IkappaB alpha phosphorylation and degradation and NF-kappaB nuclear translocation PMID: 20164171
- an IKKgamma splice variant determines IRF3 and canonical NF-kappaB pathway utilization in ssRNA virus infection PMID: 19956647
- Antioxidant amelioration of dilated cardiomyopathy caused by conditional deletion of NEMO/IKKgamma in cardiomyocytes. PMID: 19850942
- The carboxyl-terminal region of IKKgamma is required for full IKK activation PMID: 12192055
- BAFF-induced NEMO-independent processing of NF-kappa B2 in maturing B cells PMID: 12352969
- NEMO is not required for B cell development but plays an important role in B cell survival PMID: 12538858
- NEMO trimerization domain is composed of the interacting C-terminal CC2 and LZ coiled-coil subdomains PMID: 15107419
- a cell-permeable NBD peptide has been shown to block association of NEMO with the IKK complex and inhibit activation of NF-kappaB PMID: 15252035
- Activation of IKK may be mediated by direct interaction with mutant huntingtin. The expanded polyglutamine stretch & adjacent Pro-rich motifs interact with IKKgamma. Expression of IKKgamma promotes aggregation & nuclear localization of mutant Htt exon-1. PMID: 15371500
- overexpression of IKKgamma inhibits activation of NF-kappaB by NIK by competing with NIK for interaction with IKKbeta PMID: 15528996
- TLR8-mediated MEKK3-dependent IKKgamma phosphorylation might play an important role in the activation of IKK complex, leading to IkappaBalpha phosphorylation PMID: 16737960
- Deletion of the N-terminus of IKKgamma induces apoptosis in keratinocytes. PMID: 17184772
- Deletion of NEMO in liver parenchymal cells causes steatohepatitis and hepatocellular carcinoma PMID: 17292824
- protein phosphatase 2a recruitment to I-kappaB kinase gamma/NF-kappaB essential modulator is regulated by heptad repeats, which are targeted by HTLV-I tax PMID: 17314097
顯示更多
收起更多
-
亞細胞定位:Cytoplasm. Nucleus.
-
數據庫鏈接:
Most popular with customers
-
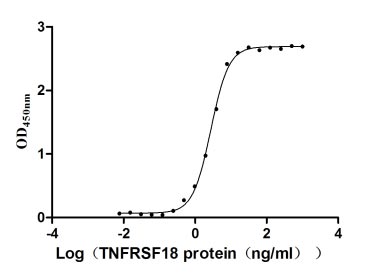
Recombinant Human Tumor necrosis factor receptor superfamily member 18 (TNFRSF18), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
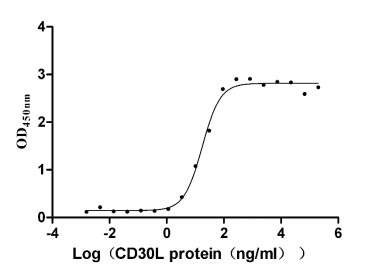
Recombinant Human Tumor necrosis factor receptor superfamily member 8 (TNFRSF8), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
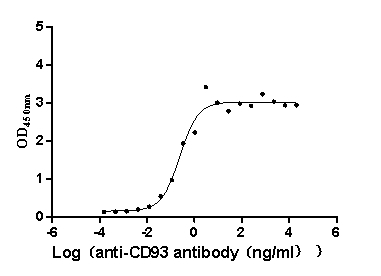
Recombinant Macaca fascicularis CD93 molecule (CD93), partial (Active)
Express system: Mammalian cell
Species: Macaca fascicularis (Crab-eating macaque) (Cynomolgus monkey)
-
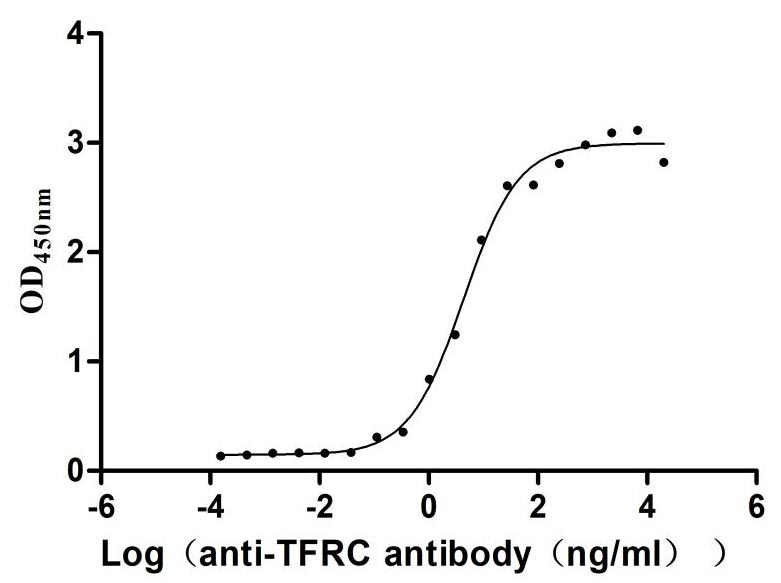
Recombinant Human Transferrin receptor protein 1 (TFRC), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
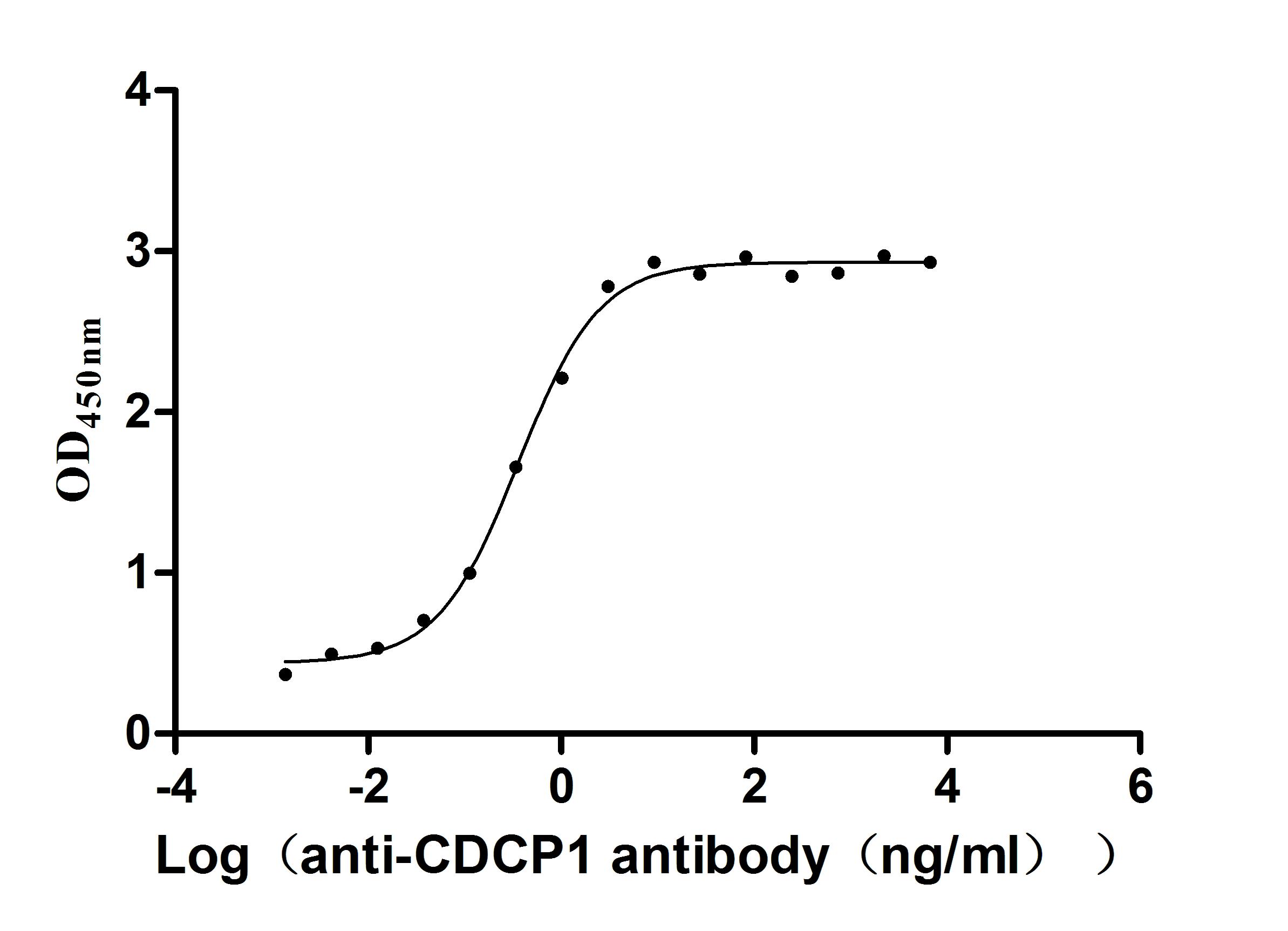
Recombinant Human CUB domain-containing protein 1 (CDCP1), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
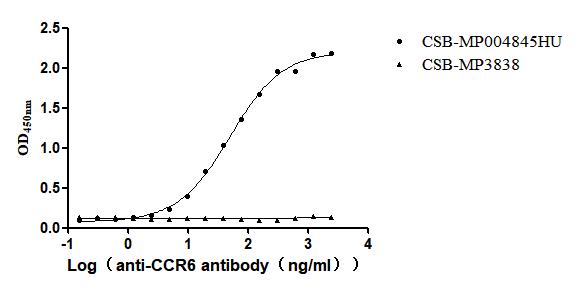
Recombinant Human C-C chemokine receptor type 6(CCR6)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
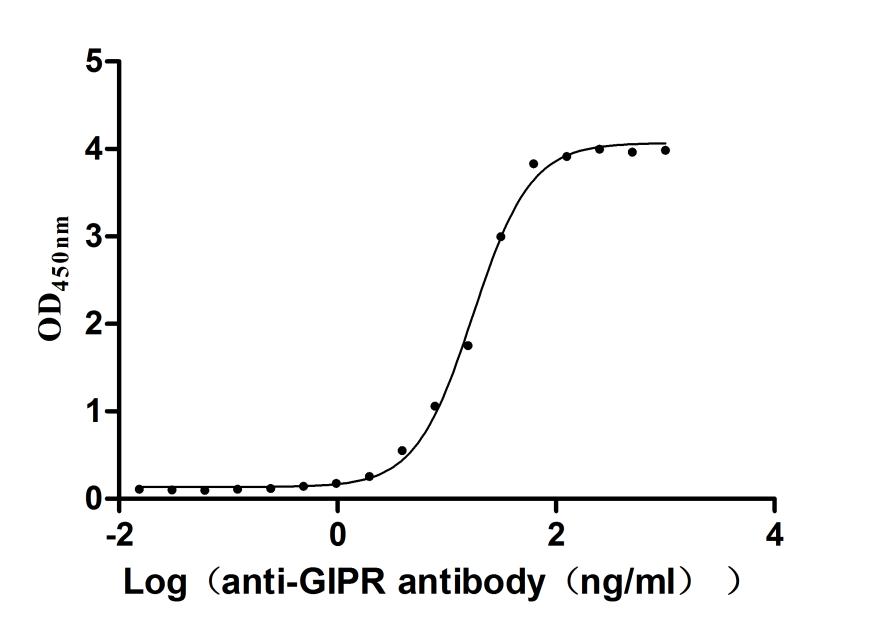
Recombinant Human Gastric inhibitory polypeptide receptor(GIPR),partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
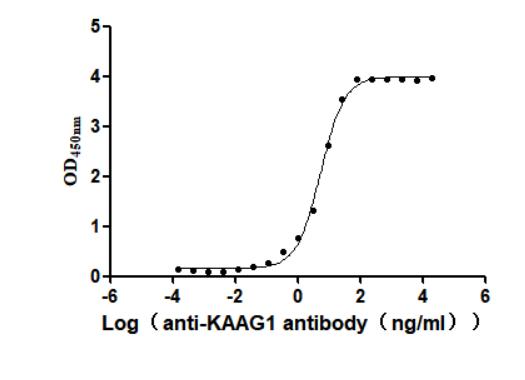
Recombinant Human Kidney-associated antigen 1(KAAG1) (Active)
Express system: Baculovirus
Species: Homo sapiens (Human)












