Recombinant Mouse Cullin-3 (Cul3)
-
中文名稱:Recombinant Mouse Cullin-3(Cul3)
-
貨號:CSB-YP881350MO
-
規格:
-
來源:Yeast
-
其他:
-
中文名稱:Recombinant Mouse Cullin-3(Cul3)
-
貨號:CSB-EP881350MO
-
規格:
-
來源:E.coli
-
其他:
-
中文名稱:Recombinant Mouse Cullin-3(Cul3)
-
貨號:CSB-EP881350MO-B
-
規格:
-
來源:E.coli
-
共軛:Avi-tag Biotinylated
E. coli biotin ligase (BirA) is highly specific in covalently attaching biotin to the 15 amino acid AviTag peptide. This recombinant protein was biotinylated in vivo by AviTag-BirA technology, which method is BriA catalyzes amide linkage between the biotin and the specific lysine of the AviTag.
-
其他:
-
中文名稱:Recombinant Mouse Cullin-3(Cul3)
-
貨號:CSB-BP881350MO
-
規格:
-
來源:Baculovirus
-
其他:
-
中文名稱:Recombinant Mouse Cullin-3(Cul3)
-
貨號:CSB-MP881350MO
-
規格:
-
來源:Mammalian cell
-
其他:
產品詳情
-
純度:>85% (SDS-PAGE)
-
基因名:
-
Uniprot No.:
-
別名:Cul3Cullin-3; CUL-3
-
種屬:Mus musculus (Mouse)
-
蛋白長度:Full Length of Mature Protein
-
表達區域:2-768
-
氨基酸序列SNLSKGTGS RKDTKMRIRA FPMTMDEKYV NSIWDLLKNA IQEIQRKNNS GLSFEELYRN AYTMVLHKHG EKLYTGLREV VTEHLINKVR EDVLNSLNNN FLQTLNQAWN DHQTAMVMIR DILMYMDRVY VQQNNVENVY NLGLIIFRDQ VVRYGCIRDH LRQTLLDMIA RERKGEVVDR GAIRNACQML MILGLEGRSV YEEDFEAPFL EMSAEFFQME SQKFLAENSA SVYIKKVEAR INEEIERVMH CLDKSTEEPI VKVVERELIS KHMKTIVEME NSGLVHMLKN GKTEDLACMY KLFSRVPNGL KTMCECMSCY LREQGKALVS EEGEGKNPVD YIQGLLDLKS RFDRFLQESF NNDRLFKQTI AGDFEYFLNL NSRSPEYLSL FIDDKLKKGV KGLTEQEVET ILDKAMVLFR FMQEKDVFER YYKQHLARRL LTNKSVSDDS EKNMISKLKT ECGCQFTSKL EGMFRDMSIS NTTMDEFRQH LQATGVSLGG VDLTVRVLTT GYWPTQSATP KCNIPPAPRH AFEIFRRFYL AKHSGRQLTL QHHMGSADLN ATFYGPVKKE DGSEVGVGGA QVTGSNTRKH ILQVSTFQMT ILMLFNNREK YTFEEIQQET DIPERELVRA LQSLACGKPT QRVLTKEPKS KEIESGHIFT VNDQFTSKLH RVKIQTVAAK QGESDPERKE TRQKVDDDRK HEIEAAIVRI MKSRKKMQHN VLVAEVTQQL KARFLPSPVV IKKRIEGLIE REYLARTPED RKVYTYVA
-
蛋白標簽:Tag?type?will?be?determined?during?the?manufacturing?process.
The tag type will be determined during production process. If you have specified tag type, please tell us and we will develop the specified tag preferentially. -
產品提供形式:Lyophilized powder
Note: We will preferentially ship the format that we have in stock, however, if you have any special requirement for the format, please remark your requirement when placing the order, we will prepare according to your demand. -
復溶:We recommend that this vial be briefly centrifuged prior to opening to bring the contents to the bottom. Please reconstitute protein in deionized sterile water to a concentration of 0.1-1.0 mg/mL.We recommend to add 5-50% of glycerol (final concentration) and aliquot for long-term storage at -20℃/-80℃. Our default final concentration of glycerol is 50%. Customers could use it as reference.
-
儲存條件:Store at -20°C/-80°C upon receipt, aliquoting is necessary for mutiple use. Avoid repeated freeze-thaw cycles.
-
保質期:The shelf life is related to many factors, storage state, buffer ingredients, storage temperature and the stability of the protein itself.
Generally, the shelf life of liquid form is 6 months at -20°C/-80°C. The shelf life of lyophilized form is 12 months at -20°C/-80°C. -
貨期:Delivery time may differ from different purchasing way or location, please kindly consult your local distributors for specific delivery time.Note: All of our proteins are default shipped with normal blue ice packs, if you request to ship with dry ice, please communicate with us in advance and extra fees will be charged.
-
注意事項:Repeated freezing and thawing is not recommended. Store working aliquots at 4°C for up to one week.
-
Datasheet :Please contact us to get it.
產品評價

靶點詳情
-
功能:Core component of multiple cullin-RING-based BCR (BTB-CUL3-RBX1) E3 ubiquitin-protein ligase complexes which mediate the ubiquitination and subsequent proteasomal degradation of target proteins. BCR complexes and ARIH1 collaborate in tandem to mediate ubiquitination of target proteins. As a scaffold protein may contribute to catalysis through positioning of the substrate and the ubiquitin-conjugating enzyme. The E3 ubiquitin-protein ligase activity of the complex is dependent on the neddylation of the cullin subunit and is inhibited by the association of the deneddylated cullin subunit with TIP120A/CAND1. The functional specificity of the BCR complex depends on the BTB domain-containing protein as the substrate recognition component. BCR(KLHL42) is involved in ubiquitination of KATNA1. BCR(SPOP) is involved in ubiquitination of BMI1/PCGF4, BRMS1, MACROH2A1 and DAXX, GLI2 and GLI3. Can also form a cullin-RING-based BCR (BTB-CUL3-RBX1) E3 ubiquitin-protein ligase complex containing homodimeric SPOPL or the heterodimer formed by SPOP and SPOPL; these complexes have lower ubiquitin ligase activity. BCR(KLHL9-KLHL13) controls the dynamic behavior of AURKB on mitotic chromosomes and thereby coordinates faithful mitotic progression and completion of cytokinesis. BCR(KLHL12) is involved in ER-Golgi transport by regulating the size of COPII coats, thereby playing a key role in collagen export, which is required for embryonic stem (ES) cells division: BCR(KLHL12) acts by mediating monoubiquitination of SEC31 (SEC31A or SEC31B). BCR(KLHL3) acts as a regulator of ion transport in the distal nephron; by mediating ubiquitination of WNK4. The BCR(KLHL20) E3 ubiquitin ligase complex is involved in interferon response and anterograde Golgi to endosome transport: it mediates both ubiquitination leading to degradation and 'Lys-33'-linked ubiquitination. The BCR(KLHL21) E3 ubiquitin ligase complex regulates localization of the chromosomal passenger complex (CPC) from chromosomes to the spindle midzone in anaphase and mediates the ubiquitination of AURKB. The BCR(KLHL22) ubiquitin ligase complex mediates monoubiquitination of PLK1, leading to PLK1 dissociation from phosphoreceptor proteins and subsequent removal from kinetochores, allowing silencing of the spindle assembly checkpoint (SAC) and chromosome segregation. The BCR(KLHL22) ubiquitin ligase complex is also responsible for the amino acid-stimulated 'Lys-48' polyubiquitination and proteasomal degradation of DEPDC5. Through the degradation of DEPDC5, releases the GATOR1 complex-mediated inhibition of the TORC1 pathway. The BCR(KLHL25) ubiquitin ligase complex is involved in translational homeostasis by mediating ubiquitination and subsequent degradation of hypophosphorylated EIF4EBP1 (4E-BP1). The BCR(KBTBD8) complex acts by mediating monoubiquitination of NOLC1 and TCOF1, leading to remodel the translational program of differentiating cells in favor of neural crest specification. Involved in ubiquitination of cyclin E and of cyclin D1 (in vitro) thus involved in regulation of G1/S transition. Involved in the ubiquitination of KEAP1, ENC1 and KLHL41. In concert with ATF2 and RBX1, promotes degradation of KAT5 thereby attenuating its ability to acetylate and activate ATM. The BCR(KCTD17) E3 ubiquitin ligase complex mediates ubiquitination and degradation of TCHP, a down-regulator of cilium assembly, thereby inducing ciliogenesis. The BCR(KLHL24) E3 ubiquitin ligase complex mediates ubiquitination of KRT14, controls KRT14 levels during keratinocytes differentiation, and is essential for skin integrity. The BCR(KLHL18) E3 ubiquitin ligase complex mediates the ubiquitination of AURKA leading to its activation at the centrosome which is required for initiating mitotic entry. The BCR(KEAP1) E3 ubiquitin ligase complex acts as a key sensor of oxidative and electrophilic stress by mediating ubiquitination and degradation of NFE2L2/NRF2, a transcription factor regulating expression of many cytoprotective genes. As part of the CUL3(KBTBD6/7) E3 ubiquitin ligase complex functions mediates 'Lys-48' ubiquitination and proteasomal degradation of TIAM1. By controlling the ubiquitination of that RAC1 guanine exchange factors (GEF), regulates RAC1 signal transduction and downstream biological processes including the organization of the cytoskeleton, cell migration and cell proliferation.
-
基因功能參考文獻:
- a loss of CULLIN3 represents a common signaling node for controlling the activity of intracellular WNT and SHH signaling pathways mediated by ID1 PMID: 27477274
- Downregulation of Cul3 led to a marked increase in RhoA protein expression after 6 days of adipocytes differentiation, suggesting that Cul3 is involved in the regulation of RhoA stability. PMID: 28499918
- Data (including data from studies using cells cultured from knockout mice) suggest that Sccro neddylates Cul3 with Nedd8, promoting Cul3/Klhl21 complex formation and localization of Cul3 during telophase. (Sccro = squamous cell carcinoma-related oncogene; Cul3 = cullin 3; Nedd8 = neural precursor cell expressed, developmentally down-regulated protein 8; Klhl21 = kelch-like protein 21) PMID: 28620047
- Results showed that in the basal state, the amount of Keap1 and Cul3 proteins were maintained at higher levels than that of Nrf2, and remained the same even under oxidative and electrophilic stimuli. PMID: 27697860
- Suggest that the hyperkalemia in knock-in mouse with the CUL3(Delta403-459) mutation is not caused by reduced ROMK expression in the distal nephron. PMID: 27378813
- These results suggest that KLHL2 likely plays a role in the pathogenesis of FHHt, and aggravates the phenotype caused by mutations in CUL3 and WNK4. PMID: 26607111
- Mutation of cul3 protein is involved in the development of renal hypertension and chronic kidney diseases. PMID: 25311533
- Data show that Bcl6 (B cell lymphoma 6)-cullin 3 complexes provide essential negative feedback regulation during both thymocyte development and T cell activation to restrain excessive T follicular helper (Tfh) responses. PMID: 24863065
- Knockdown of Cullin-3 or inhibition of cullin-RING ligase activity in aortic smooth muscle cells increased RhoA. PMID: 23040068
- CUL3 deletion in mice demonstrated an essential role for CUL3 in the development of PLZF- and BCL6-dependent lineages; distinct lineage-specific BTB-ZF transcription factors recruit CUL3 to alter the ubiquitination pattern of their associated chromatin-modifying complex PMID: 23086144
- The authors identified MUF1 as the first substrate for RhoBTB-Cul3 ubiquitin ligase complexes. PMID: 22709582
- Data show that liver-specific ablation of the cyclin E regulator Cullin 3 in mice results in the persistence and massive expansion of hepatic progenitor cells. PMID: 20978349
- Nrf2 regulates Cul3-Rbx1 by controlling regulation of expression and induction of Cul3-Rbx1 PMID: 20452971
- Results show negative regulation of HDAC1 by a Cul3-REN E3 ubiquitin ligase complex. PMID: 20081843
- Data show that the constitutive degradation pathway for cyclin E that is regulated by the Cullin3-based E3 ligase is essential to maintain quiescence in mammalian cells. PMID: 17339333
- KLHL12 specifically interacts with the D4 polymorphism, thereby building up a Cul3-E3 ligase complex with substrate specificity toward the D4 receptor PMID: 18303015
- Cul3-deficient cells or Ctb9/KLHDC5-deficient cells show an increase in p60/katanin levels, indicating that Cul3/Ctb9/KLHDC5 is required for efficient p60/katanin removal PMID: 19261606
顯示更多
收起更多
-
亞細胞定位:Nucleus. Golgi apparatus. Cell projection, cilium, flagellum. Cytoplasm. Cytoplasm, cytoskeleton, spindle. Cytoplasm. Cytoplasm, cytoskeleton, microtubule organizing center, centrosome. Cytoplasm, cytoskeleton, spindle pole.
-
蛋白家族:Cullin family
-
組織特異性:Widely expressed, with highest expression in brain, spleen and testis. In the testis, it is mainly expressed in spermatids.
-
數據庫鏈接:
Most popular with customers
-
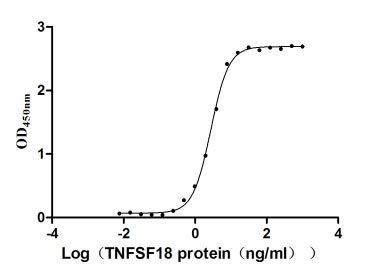
Recombinant Human Tumor necrosis factor ligand superfamily member 18 (TNFSF18), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
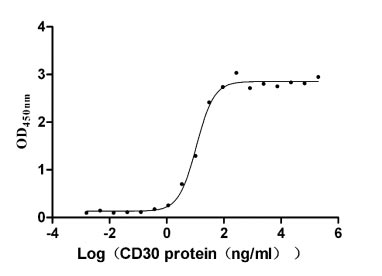
Recombinant Human Tumor necrosis factor ligand superfamily member 8 (TNFSF8), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
-AC1.jpg)
Recombinant Human T-cell surface protein tactile (CD96), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
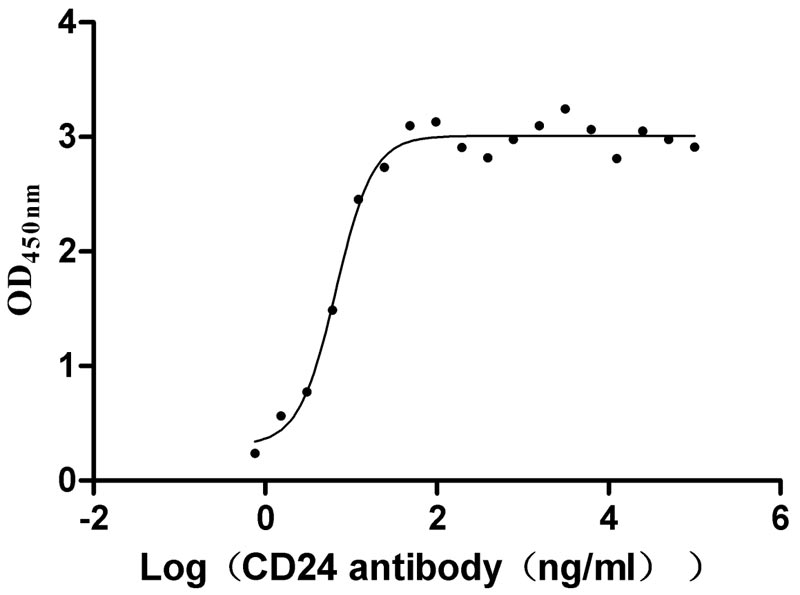
Recombinant Human Signal transducer CD24 (CD24)-Nanoparticle (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
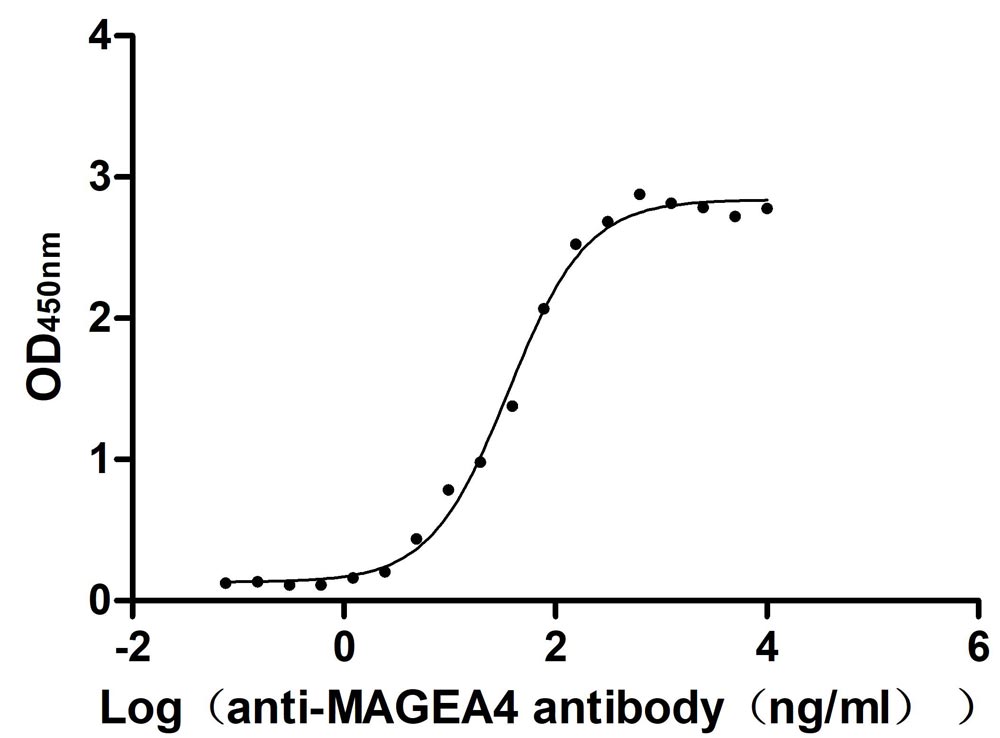
Recombinant Human Melanoma-associated antigen 4 (MAGEA4) (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Cannabinoid receptor 1 (CNR1)-VLPs (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Tomoregulin-2 (TMEFF2), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)
-
Recombinant Human Cell adhesion molecule 1 (CADM1), partial (Active)
Express system: Mammalian cell
Species: Homo sapiens (Human)












